

 首页
首页 关于我们
关于我们 产品中心
产品中心 技术服务
技术服务 技术中心
技术中心 联系我们
联系我们导读
细胞在营养物质缺乏时启动适应性反应,通过代谢途径的变化和细胞器动态调整来应对压力。在极端环境下,细胞可能会停止分裂,进入静止状态,以适应不良条件。
当细胞恢复最佳生长条件后,细胞可重新激活分裂和能量代谢。未能有效响应营养信号与人类退行性疾病、糖尿病和衰老相关,并在肿瘤微环境中有助于免疫逃逸。
在酵母中,葡萄糖饥饿限制大分子聚合物运动,导致蛋白质合成受限,而蛋白酶体和自噬活性增强,促使内质网和线粒体适应。持续的葡萄糖缺乏引起线粒体碎片化,生物能量学从糖酵解转变为呼吸作用,促进信号传导、脂肪酸氧化和清除受损线粒体。
葡萄糖缺乏初期,线粒体压力反应通过OMA1-DELE1通路传递,抑制全局蛋白质合成并激活综合应激反应(ISR)。同时,细胞质核糖体聚集在线粒体上,这是一种尚未完全理解的适应机制,显示线粒体重塑与蛋白质合成的联系,可能促进细胞器稳态和细胞存活。
2024年10月8日,美国弗吉尼亚大学Ahmad Jomaa团队在Nature Communications上发表了一篇题为“Ribosomes hibernate on mitochondria during cellular stress”的论文,阐明细胞质核糖体如何在分裂酵母中响应因葡萄糖缺乏引起的细胞压力而结合到碎片化线粒体的机制。

文章索引
【发表日期】2024年10月8日
【IF】14.7
研究结果
一. 在葡萄糖缺乏期间分离的核糖体进入非活跃状态
作者对酵母在葡萄糖缺乏条件下的蛋白质合成进行了Polysome profiling多聚核糖体分析,结果显示在低葡萄糖条件下,随着时间推移,多聚核糖体数量减少,80S单核糖体数量增加,表明蛋白质合成停止。
冷冻电镜分析显示出翻译被抑制,H69的构象变化干扰了tRNA与大亚基的结合,阻碍了蛋白质翻译的启动。

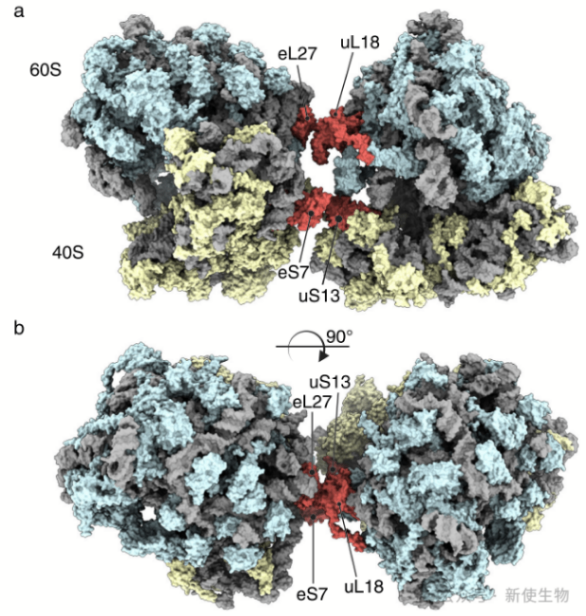
二. 细胞质中的休眠核糖体通过小核糖体亚基与线粒体结合
长期葡萄糖缺乏导致细胞质核糖体在碎片化线粒体上聚集,可能与综合应激反应(ISR)有关。
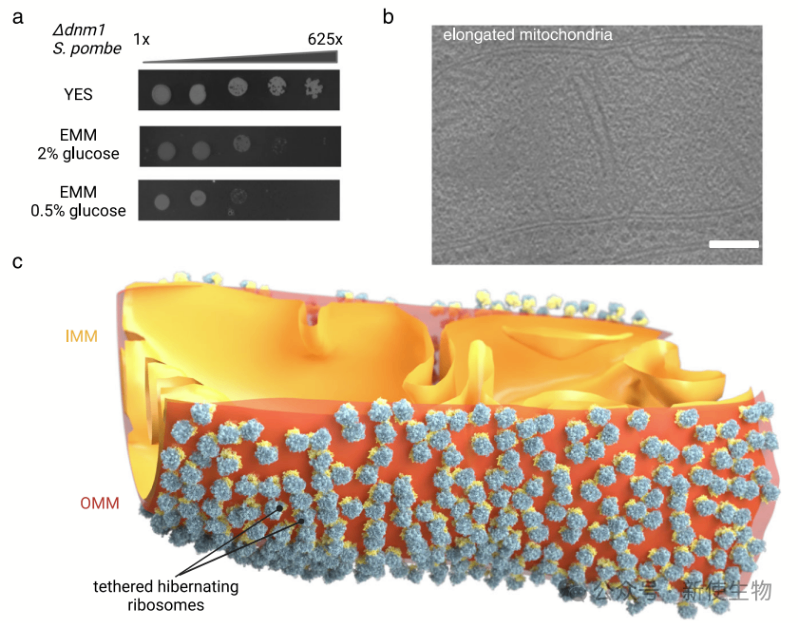
研究显示,经过2-3天细胞进入静止生长阶段,4天时线粒体出现碎片化,7天时几乎完全被核糖体覆盖。
冷冻电镜分析发现这些核糖体处于休眠状态,结合延伸因子eEF2,主要通过小亚基与线粒体结合,形成2-5个核糖体的聚集体。此机制代表了一种新型的真核生物休眠特征,可能有助于在营养恢复前保护翻译工具。

三. 核糖体的锚定与线粒体的碎片化无关
在酵母中,线粒体的碎片化由动力相关蛋白Dnm1介导,缺失Dnm1的细胞(Δdnm1)在富含培养基中表现正常,但在低葡萄糖条件下生长受限。
冷冻电镜成像显示,尽管Δdnm1细胞的线粒体保持延长状态,但在第7天时,98%的线粒体依然被核糖体覆盖。因此,核糖体锚定与线粒体碎片化无关。
四. Cpc2/RACK1介导核糖体与外膜的锚定
为探讨线粒体与休眠核糖体的相互作用,研究发现Cpc2与线粒体外膜(OMM)相距约30 Å,可能参与锚定休眠核糖体。
通过生存能力测定发现,缺失Cpc2的细胞在富营养条件下无明显生长缺陷,但在低葡萄糖培养基中显著受损。
原位冷冻电镜分析显示,尽管线粒体呈现碎片化,但缺失Cpc2的细胞在低葡萄糖条件下未观察到核糖体与线粒体的锚定,这表明Cpc2在葡萄糖缺乏条件下对核糖体锚定和细胞存活至关重要,强调其在线粒体稳态中的关键作用。

总结
本文研究发现,葡萄糖耗竭后蛋白质合成停止,核糖体缺乏tRNA和mRNA,并出现阻止tRNA结合的构象变化。核糖体蛋白Cpc2/RACK1介导核糖体与线粒体的锚定,揭示了线粒体应激与蛋白质合成停滞的分子机制,拓展了对细胞在营养缺乏下反应的理解。
超高的准确性为研究非经典的开放阅读框(ORFs)提供极大便利,提高微肽(肿瘤新生抗原)的挖掘效率。
简便快速的操作让翻译组分析不再设有门槛,每位科研学者都能轻松上手。
另外新使生物提供多物种多聚核糖体分析(Polysome profiling),了解更多翻译组技术信息可联系杭州新使生物科有限公司

感谢关注杭州新使生物,ribo seq